 вход
вход регистрация
регистрация ензими
ензими
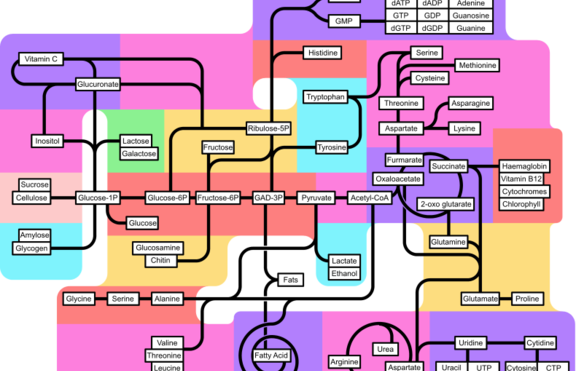
Глюконеогенезата представлява процес на синтез на глюкоза. Три са основните източници на глюкоза в клетките: лактат, пируват и глицерол. При бозайниците глюконеогенезата се осъществява предимно в черния дроб, рядко в бъбреците.
След тежко физическо натоварване лактатът (млечната киселина), който се е образувал чрез анаеробна гликолиза в мускулите, се връща в черния дроб и се преобразува в глюкоза. След това глюкозата се пренася обратно до мускулите и се съхранява под формата на гликоген (животни и човек). Това е така нареченият Цикъл на Кори.
Гликолизата (разграждане на глюкоза) и Глюконеогенезата (синтез на глюкоза) не са еднакви процеси в обратна посока, но все пак някои от стъпките се припокриват.
Седем от 10-те ензимни реакции от глюконеогенезата са точно същите като при гликолизата, но в обратна посока. Останалите три реакции в гликолизата са необратими и не биха могли да служат като път за синтез на глюкоза.
1) преобразуването на глюкозата до глюкозо-6-фосфат от хексокиназата
2) фосфорилирането на фруктозо-6-фосфат до фруктозо 1,6-бисфосфат чрез ензима фосфофруктокиназа-1
3) проебразуването на фосфоенолпирувата в пируват с ензима пируват киназа
Тези три реакции са с големи отрицателни свободни енергии и са необратими, докато останалите имат свободни енергии близки до нулата. При глюконеогенезата трите стъпки се прескачат чрез ензими, които осигуряват достатъчно енергия за това, че рекацията да бъде екзергонична. По този начин процесите гликолиза и глюконеогенеза стават необратими в клетката. Разделената регулация на двата процеса е в резултат на упражнявания ензимен контрол, който е уникален, както за гликолизата, така и за глюконеогенезата.
Превръщането на пируват във фосфоенолпитуват се осъществява чрез две реакции, които са екзергонични. Пируватът се транспортира до митохондриите след гликолизата (може да е получен и в самите митохондрии от аланин). Ензимът пируват карбоксилаза чрез своя коензим биотин преобразува пирувата в оксалацетат. Участва една молекула АТФ. Тъй като митохондиралната мембрана няма транспортери за оксалацетат, за да се пренесе в цитозола, той се редуцира до малат чрез малат дехидрогеназа с разход на NADH.
След като се получи малатът, се пренася в цитозола със специални транспортери и отново се превръща в оксалацетат, при което NADH се възвръща. Ензимът фосфоенолпируват карбоксилаза катализира преобразуването на оксалацетата във фосфоенолпируват. При тази реакция молекула ГТФ е донов на фосфорилна група.
Другият начин пируватът да се превърне във фосфоенолпируват е ако лактат е източник. Този път позволява лактатът, който се е синтезирал при гликолизата в еритроцитите например или в мускулите, да бъде използван за синтез на глюкоза. Това е един от най-често използваните пътища са синтез на глюкоза при гръбначните животни и хора след тежко физическо натоварване. При проебразуването на лактат до пируват в цитозола има добив на NADH и следователно малатът от митохондриите не е необходим. След като пируватът, получен от лактат дехидрогеназната реакция се пренесе в митохондиите, се превръща в оксалацетат чрез ензима пируват карбоксилаза. След това оксалацетатът се превръща във фосфоенолпируват чрез изоензима фосфоенолпируват карбоксилаза и се изнася в цитозола, за да се продължи глюконеогенезата. Двата ензима фосфоенолпируват карбоксилаза (единият в цитозола, а другият в митохондриите) се кодират от различни гени в хромозомите, позволявайки два ензима да катализират една и съща реакция, но в различни места в клетката, както и да имат различни роли (изоензими).
Другата рекация - преобразуването на 1,6-бисфосфат до фруткозо-6-фосфат се катализира от ензима фруктозо-1,6-бисфосфатаза.
Преобразуването на глюкозо-6-фосфат в глюкоза е последната стъпка от глюконеогенезата. При реакцията, катализирана от флюкозо-6-фосфатаза не се получава АТФ, тя е проста хидролиза на фосфатния естер. Магнезиеви йони активират ензима.
За образуването на всяка молекула глюкоза се използват 6 високо-енергийни фосфатни групи, четири от АТФ и две от ГДФ. Две молекули NADH са използвани при редукцията на 1,3-бисфосфоглицерат.
Глюкозата може да бъде получена и от междинни метаболити на цикъла на Кребс, които могат да се окислят до оксалацетат. Аминокиселините също могат да служат са синтеза на глюкоза. Аланин и Глутамин са от особено значение за бозайниците като глюкогенни аминикиселини.
Гликолизата и Глюконеогенезата се регулират реципрочно. Регулират се алостерично и чрез ковалентни модификации (фосфорилиране). Когато потокът на гликолизата се покачи, потокът на глюконеогенезата спада и обратно.
Образуването на една молекула глюкоза е доста "скъп" процес: изразходват се 4АТФ молекули, 2ГТФ и 2NADH.











 Как всяка зодия става по-уверена с навлизането на Марс в Лъв
Как всяка зодия става по-уверена с навлизането на Марс в Лъв  Украинският лазер Tryzub може да унищожи руски дронове
Украинският лазер Tryzub може да унищожи руски дронове  Новият боен кораб на Северна Корея е най-големият в нейната история
Новият боен кораб на Северна Корея е най-големият в нейната история  Линда Еванджелиста продава своя красив пентхаус в Ню Йорк - вижте го
Линда Еванджелиста продава своя красив пентхаус в Ню Йорк - вижте го  „България Еър“ стартира новата линия Варна – Париж - Варна
„България Еър“ стартира новата линия Варна – Париж - Варна  Сбогом, зимни обувки – маратонките, които ще носим през пролетта
Сбогом, зимни обувки – маратонките, които ще носим през пролетта  22 години от премиерата на филма на Джулия Робъртс „Усмивката на Мона Лиза“
22 години от премиерата на филма на Джулия Робъртс „Усмивката на Мона Лиза“  Деми Мур знаела, че ще загуби надпреварата за „Оскар“
Деми Мур знаела, че ще загуби надпреварата за „Оскар“  „България Еър“ стартира новата линия Варна – Париж – Варна
„България Еър“ стартира новата линия Варна – Париж – Варна  4 вкусни рецепти за дроб сарма за Великден
4 вкусни рецепти за дроб сарма за Великден  Поверия за Разпети петък
Поверия за Разпети петък  Три зодии трябва да са внимателни по време на Меркурий в Овен
Три зодии трябва да са внимателни по време на Меркурий в Овен  Космическите отпадъци достигнаха нов тревожен рекорд
Космическите отпадъци достигнаха нов тревожен рекорд  Кристалина Георгиева: Държавите трябва да извлекат най-доброто от новия многополюсен свят
Кристалина Георгиева: Държавите трябва да извлекат най-доброто от новия многополюсен свят  Късна емисия
Късна емисия  дава под наем, Двустаен апартамент, 55 m2 София, Студентски Град, 550 EUR
дава под наем, Двустаен апартамент, 55 m2 София, Студентски Град, 550 EUR  дава под наем, Офис, 530 m2 София, Гео Милев, 3445 EUR
дава под наем, Офис, 530 m2 София, Гео Милев, 3445 EUR  дава под наем, Офис, 737 m2 София, Гео Милев, 4791 EUR
дава под наем, Офис, 737 m2 София, Гео Милев, 4791 EUR  дава под наем, Офис, 340 m2 София, Гео Милев, 2178 EUR
дава под наем, Офис, 340 m2 София, Гео Милев, 2178 EUR